
| Taxa: |
| Order: |
| Family: |
| Mammalia |
| Chiroptera |
| Vespertilionidae |
| NatureServe Global Rank: |
| NatureServe State (NC) Rank: |
| G4 |
| SUB,S2N |
| Federal Status: |
| NC State Status: |
| --- |
| SC |
| Land Unit |
| US Fish & Wildlife Service |
| US Forest Service |
| US National Park Service |
| US Department of Defense |
| NC State Parks |
| NC University System |
| NC Wildlife Res. Com. |
| NC Forest Service |
| NC Div. of Coastal Mgmt. |
| Local Governments |
| Non-Governmental Org. |
| Other Public Lands |
| Private Lands |
| GAP Status 1-2 |
| All Protected Lands |
| Statewide |
| Hectares |
| 0.00 |
| 332,062.38 |
| 124.83 |
| 96,991.02 |
| 3,835.80 |
| 26.01 |
| 14,554.44 |
| 2,749.17 |
| 0.00 |
| 7,606.98 |
| 5,954.13 |
| 1.35 |
| 995,654.52 |
| 152,393.88 |
| 463,175.13 |
| 1,459,560.63 |
| Acres |
| 0.00 |
| 820,543.85 |
| 308.46 |
| 239,669.98 |
| 9,478.47 |
| 64.27 |
| 35,964.80 |
| 10,345.14 |
| 0.00 |
| 18,797.25 |
| 14,712.97 |
| 3.34 |
| 2,460,315.42 |
| 380,125.20 |
| 1,148,082.24 |
| 3,610,203.95 |
| % of Dist. on |
| Prot. Lands |
| 0.0 % |
| 71.7 % |
| 0.0 % |
| 20.9 % |
| 0.8 % |
| < 0.1 % |
| 3.1 % |
| 0.6 % |
| 0.0 % |
| 1.2 % |
| 1.2 % |
| < 0.1 % |
| 0.0 % |
| 32.9 % |
| ----- |
| ----- |
| % of Dist. on |
| All Lands |
| 0.0 % |
| 22.8 % |
| < 0.1 % |
| 6.6 % |
| 0.3 % |
| < 0.1 % |
| 1.0 % |
| 0.2 % |
| 0.0 % |
| 0.5 % |
| 0.4 % |
| < 0.1 % |
| 68.2 % |
| 10.4 % |
| ----- |
| ----- |
|
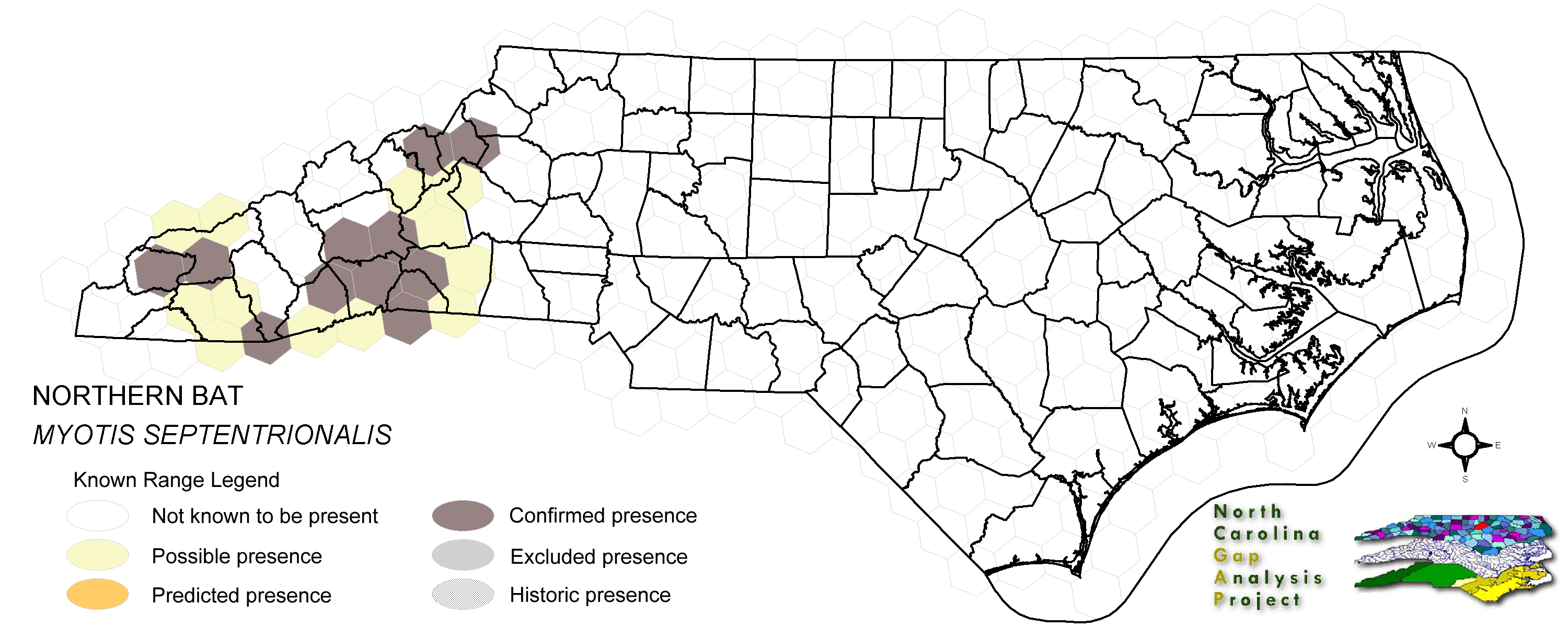
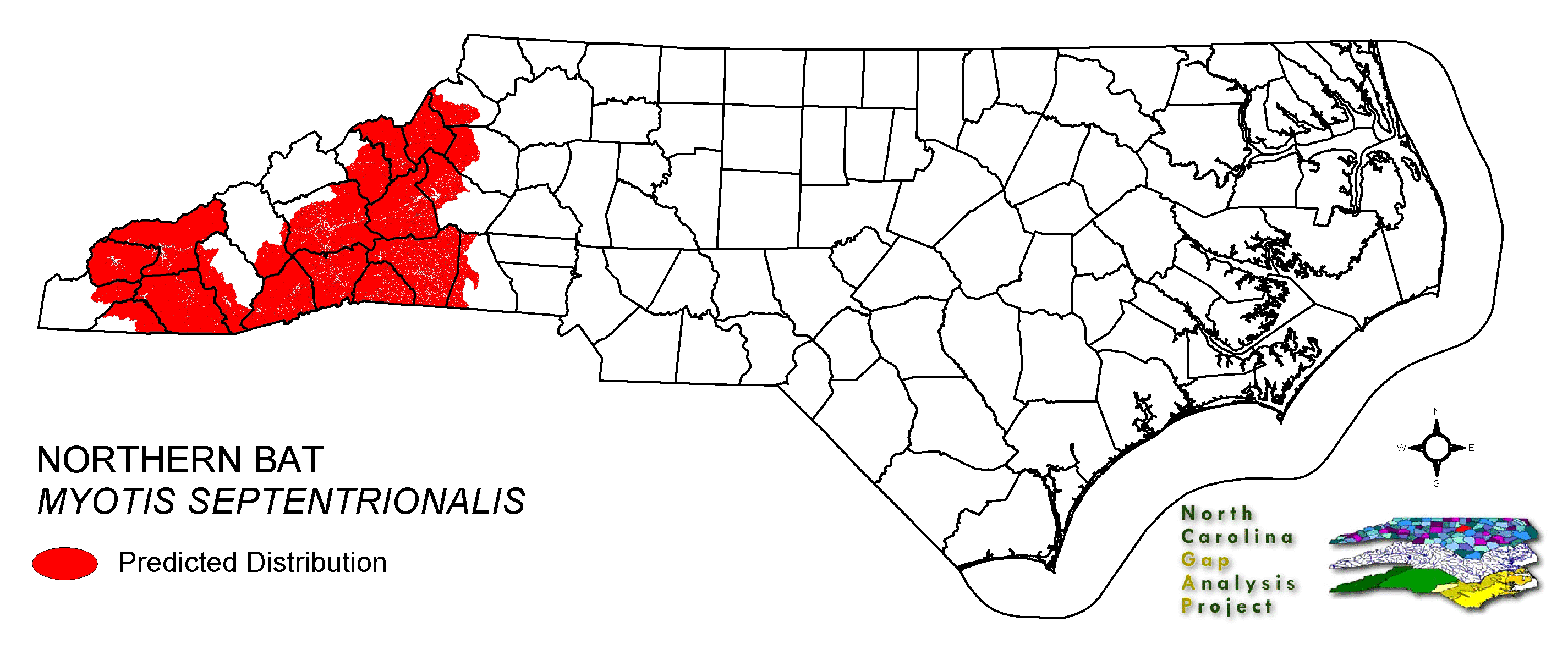
This bat is an uncommon resident of the mountains and piedmont of North Carolina (Webster et al. 1985). Often solitary or in small groups, these bats will roost in a variety of partially to fully concealed sites. Crevices in tree bark and exterior 'nooks and crannies' of buildings appear to be all that the bat requires for secondary (or temporary) roosts (Whitaker and Hamilton 1998). Caves or deeper recesses of buildings may be most used as primary or recurrent roost sites (Webster et al. 1985). From few maternity colony records, small groups of females locate in hollow trees, crevices in tree bark, bat houses and other manmade structures such as barns and little-used warehouses (Whitaker and Hamilton 1998). NATURE SERVE GLOBAL HABITAT COMMENTS: Generally associated with forested communities. Hibernates in caves, mines, and tunnels from late fall through early spring (Kurta 1982, Mumford and Cope 1964, Jackson 1961, Griffin 1940, and others). Hibernators frequently roost in crevices, drill holes, and similar sites (Caire et al. 1979, Pearson 1962, Layne 1958, Griffin 1940), but roosting in the open is not uncommon. Use of different types of hibernacula can vary considerably among areas, depending upon quality and availability of sites. In a study of 71 potential hibernation sites, including large and small caves, overhangs, and mines, on the Shawnee National Forest in southern Illinois (Whitaker and Winter 1977), mines were the only occupied habitat. Mines also are the principal hibernation sites in northern Michigan where there are no caves (Stones 1981). In the northeastern U.S., hibernation sites include mines and caves (Griffin 1940, Hall et al. 1957) as well as large, cavelike water conduit tunnels (G. Hammerson, pers. obs.). The principal requirements of a suitable hibernation site are winter-long, low temperatures above freezing, high humidity, and lack of disturbances, both natural (floods) and anthropogenic (visitation) (Barbour and Davis 1969, Hitchcock 1949). At least two studies have provided contradictory information on thermal habitat preferences, suggesting that warmer temperatures sometimes are selected or at least tolerated. In Illinois, Pearson (1962) found that the mean temperature at hibernation sites averaged 9.7 C. Stones (1981) studied the occurrence of bats in northern Michigan mines that were vertically thermally stratified. The mean ambient temperature was 5.9 C, with 43% of the population occurring in the range 7-8 C and 6.5% occurring in the range 9-11 C. There appears to be a high degree of philopatry in hibernaculum use. In Missouri, over 90% of recaptured banded individuals, representing 5% of the original banded population of 945 (753 males and 192 females), were recaptured at their cave of origin (Caire et al. 1979). Mills (1971) recaptured 4.8% of 358 individuals at their cave of origin the year after banding. Griffin (1945) found that of over 13,000 banded bats of various species, of which about 8,500 were banded in their winter hibernacula, the ratio was 100:1 for bats that were observed to return to their cave of origin over subsequent winters vs. those that were recaptured elsewhere. Night roosts used in summer between foraging bouts are in different habitats than day roosts. Caves, mines, and quarry tunnels are used as night roosts, typically by males, but also by nonreproductive females (Clark et al. 1987, Jones et al. 1967). They are joined later in the summer by juveniles and post-lactating females (Kunz, 1973). During the day, these same sites usually house no M. SEPTENTRIONALIS. Daytime observations typically are of individuals under loose bark on trees and in a variety of small spaces associated with buildings and other structures (Hoffmeister 1989, Caire et al. 1979, Hamilton and Whitaker 1979, Barbour and Davis 1969). At times M. SEPTENTRIONALIS has been found in or around caves on summer nights, but not actually roosting in them (Mills 1971). Early in the summer, these groups mostly comprise males, with females and young-of-the-year joining later in the season (Caire et al. 1979). Copulation occurs in the fall during the swarming period when large numbers of bats congregate in and near certain caves (Baker 1983, Kurta 1980). Nothing has been published on the fidelity of individuals or colonies to particular swarming sites, nor the relationship of swarming site selection to hibernaculum and summer roost selection. Given the low numbers found in most hibernacula and summer night roosts relative to the higher numbers found at swarming sites, it appears that certain caves serve as congregation points for fall mating activity. However, short-term banding returns at swarming sites are very low, indicating movement among swarming sites (Kurta, pers. comm.). Nursery colonies have rarely been located. Those that have been found were small and in a variety of sites, including a barn (Cope and Humphrey 1972) and a small cabin (Brandon 1961), though most likely the majority occur under the loose bark of trees, similar to the colonies reported from Indiana (Mumford and Cope 1964). Perhaps the single largest colony reported was found in a barn in Indiana (Cope and Humphrey 1972) on 22 June comprising 24 adult females, 12 immature females, and 18 immature males; 10 other adults escaped. Of the 24 females, 23 were lactating and 1 was pregnant. The ratio of number of adults to young confirmed the suspected litter size of one. Maternity roosts are warm sites that maximize the growth rate of young while providing protection from predation and the weather. Cool summer temperatures can slow juvenile growth, thereby reducing the fat accumulation period prior to hibernation, and ultimately increasing the risk of overwintering mortality in young-of-the-year (McNab 1982). Sex ratios from summer studies in different habitats demonstrate sexual dimorphism in habitat selection at this time of the year. In riparian areas in Iowa, Kunz (1971) found a 2:1 ratio of females to males, with most males taken in May and late August, apparently during migration. Caire et al. (1979), trapping at caves, found a preponderance of males. Small, highly fragmented, or young forests that provide limited areas of subcanopy foraging habitat may not be suitable. Young forests may also lack appropriate nursery sites. A lack of suitable hibernacula may prevent occupancy of areas that otherwise have adequate habitat (Kurta 1982). |
| Code | Name | Description | NC Natural Heritage Program Equivalent |
| 238 | Piedmont/Mountain Submerged Aquatic Vegetation | Seasonally to permanently flooded areas with aquatic vegetation. Waterlily, pondweed, hydrilla smartweed are a few of the species that can occur. | Piedmont/Mountain Semipermanent Impoundment (in part) |
| 239 | Piedmont/Mountain Emergent Vegetation | Emergent vegetation of all wetland hydrologies. Sites would commonly support species such as tussock sedge, rushs, and cattail alliances. | Rocky Bar and Shore (in part) |
| 267 | Riverbank Shrublands | Riverside shrubs with temporarily flooded hydrologies. Found in the both the Mountains and Piedmont. Containing dominants such as smooth alder and a Carolina or black willows. | Sand and Mud Bar |
| 269 | Floodplain Wet Shrublands | Saturated shrublands of the Piedmont, includes buttonbush, swamp-loosestrife, decodon and alders. | Piedmont/mountain Semipermanent Impoundment |
| 230 | Piedmont Mesic Forest | American Beech - Red Oak - White Oak Forests. | Mesic Mixed Hardwood |
| 384 | Piedmont/Mountain Mixed Bottomland Hardwood Forests | Includes temporarily to seasonally forests dominated by hardwood species. Hardwoods include sweetgum, red maple, sycamore which co-occur in a mosaic of bottomland and levee positions. Includes alluvial hardwood forests in the mountains. Hemlock and white pine may occur as inclusions, but are generally mapped separately. | Piedmont/Mountain Alluvial Forest, Piedmont/Mountain Levee Forest |
| 383 | Piedmont Mixed Successional Forest | Generally loblolly mixed with successional hardwoods. Sweetgum, tulip poplar and red maple are common co-dominants in these successional forests. | No equivalent |
| 228 | Piedmont Dry-Mesic Oak and Hardwood Forests | Primarily oak dominated forests, white oak is often dominant, with co-dominants including . Also represented by sweetgum and tulip poplar dominated forests. | Dry Mesic Oak Hickory Forest, Basic Oak Hickory Forest, Dry Oak Hickory Forest |
| 222 | Piedmont Dry-Mesic Pine Forests | Loblolly dominated forests resulting from succession following clearing. This type occurs on all moisture regimes following disturbance with the exception of the extremely xeric sites. | No equivalent |
| 382 | Dry Mesic Oak Pine Forests | Mixed forests of the coastal plain and piedmont. Includes loblolly pine with white, southern red and/or post oak and loblolly with water oak. On basic sites of the piedmont, eastern red cedar may co-occur with post, black, and blackjack oaks. | Dry Mesic Oak Hickory Forest, Xeric Hard Pan Forest, Chestnut Oak Forest, Dry Mesic Oak Hickory Forest, Dry Oak Hickory Forest |
| 220 | Piedmont Xeric Pine Forests | Dry to xeric pine forests dominated by Virginia pine, shortleaf pine or Eastern Red Cedar. | Pine Oak Heath |
| 226 | Piedmont Xeric Woodlands | Generally post and blackjack oak dominated woodlands. White ash and pignut hickory can be found in combination with Eastern red cedar on glades. | Xeric Hardpan Forest |
| 20 | Coniferous Regeneration | Regenerating pine stands. Predominantly loblolly pine, but slash and longleaf stands occur as well. | No equivalent |
| 21 | Coniferous Cultivated Plantation (natural / planted) | Managed pine plantations, densely planted. Most planted stands are loblolly, but slash and longleaf occur as well. | No equivalent |
| 51 | Deciduous Cultivated Plantation | Planted deciduous trees. Includes sweetgum and sycamore plantations. | No equivalent |
| 36 | Successional Deciduous Forests | Regenerating deciduous trees with a shrub stature. Commonly dominated by sweetgum, tulip poplars and maples. | No equivalent |
| 180 | Agricultural Crop Fields | Farm fields used for row crops. | No equivalent |
| 205 | Agricultural Pasture/Hay and Natural Herbaceous | Farm fields used for pasture grass or hay production, as well as old fields dominated by native and exotic grasses. | No equivalent |
| 202 | Residential Urban | Includes vegetation interspersed in residential areas. Includes lawns, mixed species woodlots, and horticultural shrubs. Vegetation accounts for between 20 - 70% of the cover. | No equivalent |
| 203 | Urban Low-Intensity Developed | Highly developed areas with vegetation accounting for < 20% of the cover. | No equivalent |
| 517 | Hemlock Floodplain Forest | Alluvial forest with hemlock and/or white pine in mountains and western piedmont. Hydrology is generally temporarily to seasonally flooded. | Canada Hemlock Forest |
| 521 | Spruce/Fir Forest | High Elevation Frazer-Fir - Red Spruce, Red Spruce and Red-Spruce-Yellow Birch Forests. Tree densities included here include both woodland to forest density. Highly intermixed with Northern Hardwoods, Grassy Balds, and Shrub Balds. | Red Spruce--Fraser Fir Forest, Fraser Fir Forest |
| 522 | Northern Hardwoods | High Elevation forests including yellow birch, American beech, and yellow buckeye. Includes forests with Hemlock and Yellow Birch. | Northern Hardwoods Forest, Boulderfield Forest |
| 523 | Grassy Bald | High Elevation grassy balds including Pennsylvania sedge, mountain oatgrass, as well as shrubby areas dominated by Alleghany and smooth blackberry. | Grassy Bald |
| 524 | Shrub Bald | Variable phenologies, predominantly evergreen balds with rhododendon and Mountain laurels. Deciduous shrubs including green alder and Alleghany and smooth blackberry are included as well. Red Oak - Chestnut Oak Woodlands may be included in cases where the density of the woodland species is low and the shrub component is dense. | Heath Bald |
| 525 | Appalachian Oak Forest | A variety of oak forest types including Black, White, Scarlet Oaks in dry to mesic situations. Includes forests historically co-dominated by American Chestnut. | High Elevation Red Oak Forest, Montane White Oak Forest |
| 526 | Appalachian Cove Forest | Mixed Mesophytic forests of the mountains. Includes tuliptree, basswood, yellow buckeye and surgar maple. This class is mapped to include cove forests dominated or co-dominated by hemlock. | Rich Cove Forest, Acidic Cove Forest |
| 527 | Appalachian Hemlock | Upland hemlock forests of the moutains region. Vary from side slopes to steep slope positions. | Canada Hemlock Forest |
| 528 | Appalachian Xeric Pine Forest | Pine forests and woodlands on xeric sites. A variety of pines, including Virginia, Shortleaf, Eastern White Pine, Table Mountain and Pitch pine. Often small areas of dense pine within a matrix of Xeric Oak-Pine Forests. | Pine Oak Heath |
| 529 | Appalachian Xeric Mixed Forest | Mixed forests with Virginia, Shortleaf, Eastern White Pine, Table Mountain and Pitch pines in combination with xeric oak species. Oaks include, white, Southern Red, black, and rock chestnut. | Pine Oak Heath |
| 530 | Appalachian Xeric Deciduous Forest | Deciduous forests in the mountains dominated by Xeric Oak species. Species include, white, Southern red, black, and rock chestnut. | High Elevation Red Oak Forest, Montane White Oak Forest |
| 533 | Appalachian Swamp Forest | Evergreen and deciduous forests with saturated hydrologies. This class may contain a variety of trees species, including hemlock - red maple, pitch pine, and white pine forests. | Swamp Forest-Bog Complex, Southern Appalachian Bog, Southern Appalachian Fen |
| 534 | Appalachian Wet Shrubland/ Herbaceous | Saturated shrubs and herbaceous vegetation. Often mapped as an inclusion in Appalachian Swamp Forest. | Southern Appalachian Bog, Southern Appalachian Fen |
| 535 | Talus/Outcrops/Cliffs | Includes seep talus slopes with sparce vegetation, as well as outcrops including, granitic outcrops. Some outcrops will have been mapped as barren rock. | No equivalent |
|
Guthrie, M. J. 1933. The reproductive cycles of some cave bats. Journal of Mammalogy 14:199-216.
Griffin, D. R. 1940. Notes on the life histories of New England cave bats. Journal of Mammalogy 21:181-7. Griffin, D.R. 1940. MIGRATIONS OF NEW ENGLAND BATS. BULL. MUS. COMP. ZOOL. 86(6):217-246. LaVal, R. K., R. L. Clawson, M. L. LaVal, and W. Caire. 1977. Foraging behavior and nocturnal activity patterns of Missouri bats, with emphasis on the endangered species Myotis grisescens and Myotis sodalis. J. Mamm. 58:592-599. Whitaker, J. O. Jr, and F. A. Winter. 1977. Bats of the caves and mines of the Shawnee National Forest, southern Illinois. Ill. State Acad. Sci. 70(3/4):301-13. van Zyll de Jong, C. G. 1979. Distribution and systematic relationships of long-eared MYOTIS in western Canada. Canadian J. Zool., 57:987-994. Kurta, A. 1980. Notes on summer bat activity at Michigan caves. Natl. Speleolog. Soc. Bull. 42:68-9. Fenton, M. B. 1982. Echolocation, insect hearing, and feeding ecology of insectivorous bats. Pages 261-85 in T. H. Kunz (editor). Ecology of Bats. Plenum Press, New York, New York. Kurta, A. 1982. A review of Michigan Bats:Seasonal and geographic distribution. Mich. Acad. 14(3):295-312. McNab, B. K. 1982. Evolutionary alternatives in the physiological ecology of bats. Pages 151-200 in T. H. Kunz (editor). Ecology of Bats. Plenum Press, New York, New York. Racey, P. A. 1982. Ecology of bat reproduction. Pages 57-104 in T. H. Kunz (editor). Ecology of Bats. Plenum Press, New York, New York. Jones, J. K., Jr., et al. 1992. Revised checklist of North American mammals north of Mexico, 1991. Occas. Pap. Mus., Texas Tech Univ. (146):1-23. Hammerson, Geoffrey. Research Zoologist, The Nature Conservancy, 68 Landing Road, Higganum, CT 06441. Personal communication. Stones, R. C. 1981. Survey of winter bat populations in search of the Indiana bat in the western Upper Peninsula of Michigan. Report submitted to Michigan Department of Natural Resources. 20 pp. Caire, W., R. K. LaVal, M. L. LaVal, and R. Clawson. 1979. Notes on the ecology of MYOTIS KEENIII (Chiroptera, Vespertilionidae) in Eastern Missouri. Amer. Midl. Nat. 102(2):404-7. Whitaker, J. O., Jr., and L. J. Rissler. 1992. Winter activity of bats at a mine entrance in Vermillion County, Indiana. Am. Midl. Nat. 127:52-59. Wilson, D. E., and D. M. Reeder (editors). 1993. Mammal Species of the World:a Taxonomic and Geographic Reference. Second Edition. Smithsonian Institution Press, Washington, DC. xviii + 1206 pp. Griffin, D.R. 1945. TRAVELS OF BANDED CAVE BATS. J. MAMMAL. 26:15-23. Hitchcock, H. B. 1949. Hibernation of bats in southeastern Ontario and adjacent Quebec. The Canadian Field Naturalist 63(2):47-59. Whitaker, J.O. Jr. and W.J. Hamilton, Jr. 1998. Mammals of the eastern United States. Cornell Univ. Press, Ithaca, New York. 583 pp. Hall, J. S., R. J. Cloutier, and D. R. Griffin. 1957. Longevity records and notes on tooth wear of bats. Journal of Mamalogy 38:407-409. Layne, J. N. 1958. Notes on animals of southern Illinois. Amer. Midl. Nat. 60(1):219-54. Jones, J. K. Jr., E. D. Fleharty, and P. B. Dunnigan. 1967. The distributional status of bats in Kansas. Univ. Kans, Mus. Nat. Hist. Misc. Pub. 46:1-33. Kurta, A. Biology Department. Eastern Michigan University, Ypsilanti, Michigan 48197. Personal communication. LaVal, R. K., and M. L. LaVal. 1980. Ecological studies and management of Missouri bats, with emphasis on cave dwelling species. Terrestrial Series 8, Missouri Department of Conservation. Jackson, H.H. 1961. Mammals of Wisconsin. Univ. Wisc. Press, Madison. 504 pp. Brandon, R. A. 1961. Observations of young keen bats. Journal of Mammalogy 42(3):400-1. Pearson, E. W. 1962. Bats hibernating in silica mines in southern Illinois. Journal of Mammalogy 43(1):27-33. Barbour, R. W., and W. H. Davis. 1969. Bats of America. The University of Kentucky Press, Lexington, Kentucky. Mumford, R.E. and J.B. Cope. 1964. Distribution & status of the Chiroptera of Indiana. Am. Midl. NAt. 72(2):473-489. Easterla, D. A. 1965. Parturition of Keen's myotis in southwestern Missouri. Journal of Mammalogy 49(4):770. Hitchcock, H. B., R. Keen, and A. Kurta. 1984. Survival rates of MYOTIS LEIBII and EPTESICUS FUSCUS in southeastern Ontario. Journal of Mammalogy 65:126-30. Layne, J. N., editor. 1978. Rare and endangered biota of Florida. Vol. 1. Mammals. State of Florida Game and Freshwater Fish Commission. xx + 52 pp. Hamilton, William J., Jr., and John O. Whitaker, Jr. 1979. Mammals of the eastern United States. Cornell Univ. Press, Ithaca, New York. 346 pp. Griffith, L. A., and J. E. Gates. 1985. Food habits of cave-dwelling bats in the central Appalachians, Journal of Mammalogy 66(3):451-60. Hall, E. R. 1981. The Mammals of North America. Second edition. 2 Volumes. John Wiley and Sons, New York, New York. Baker, Rollin H. 1983. Michigan mammals. Michigan State University Press. 642 pp. van Zyll de Jong, C. G. 1985. Handbook of Canadian Mammals. Volume 2. Bats. National Museums of Canada, Ottawa, Ontario, Canada. 212 pp. Webster, W. D., J. F. Parnell and W. C. Biggs Jr. 1985. Mammals of the Carolinas, Virginia, and Maryland. The University of North Carolina Press, Chapel Hill, NC. Clark, B. K., J. B. Bowles, and B. S. Clark. 1987. Summer status of the endangered Indiana bat in Iowa. Amer. Midl. Nat. 118(1):32-9. Fenton, M. B. 1969. Summer activity of MYOTIS LUCIFUGUS (Chiroptera:Vespertilionidae) at hibernacula in Ontario and Quebec. Can. J. Zool. 47:597-602. Hoffmeister, D. F. 1989. Mammals of Illinois. University of Illinois Press. 349 pp. Stones, R. C., and L. P. Branick. 1969. Use of hearing in homing by two species of MYOTIS bats. Journal of Mammalogy 50(1):157-60. Kunz, T. H. 1971. Reproduction of some vespertilionid bats of central Iowa. Amer. Midl. Nat. 86(2):477-86. Mills, R. S. 1971. A concentration of MYOTIS KEENII at caves in Ohio. Journal of Mammalogy 52:625. Whitaker, J. O. Jr., and R. E. Mumford. 1971. Notes on a collection of bats taken by mist-netting at an Indiana cave. Amer. Midl. Nat. 85(1):277-9. Cope, J. B., and S. R. Humphrey. 1972. Reproduction of the bats MYOTIS KEENII and PIPISTRELLUS SUBFLAVUS in Indiana. Bat Res. News 13:9-10. Whitaker, J. O. Jr. 1972. Food habits of bats from Indiana. Can. J. Zoology. 50:877-83. Kunz, T. H. 1973. Resource utilization:Temporal and spatial components of bat activity in cental Iowa. Journal of Mammalogy 54(1):14-32. |
For more information please contact them at:
NC-GAP Analysis Project
Dept. of Zoology, NCSU
Campus Box 7617
Raleigh, NC 27695-7617
(919) 513-2853
www.basic.ncsu.edu/ncgap